Amidon
Composition et propriétés
L'amidon est le polysaccharide de réserve le plus répandu chez les plantes.
L'amidon est un mélange de deux α-D-glucanes : l'amylose et l'amylopectine. La teneur de l'amidon en ces deux types moléculaires varie avec l'espèce végétale. L'amylose est en général le constituant le moins abondant de l'amidon. Les amidons de blé et de pomme de terre qui en sont les plus riches n'en contiennent que de 20 à 30%.
Dans les cellules végétales, l'amidon est stocké dans des particules insolubles partiellement cristallines appelées grains d'amidon, dont les diamètres varient de 1 à 100 micromètres. On trouve ces granules dans les chloroplastes, où l'amidon est synthétisé, et dans les amyloplastes qui en constituent le réservoir à long terme. Les granules présentent une structure feuilletée, formée de zones concentriques de 100-400 nm d'épaisseur, responsables du phénomène bien connu de la croix noire à l'observation microscopique en lumière polarisée.
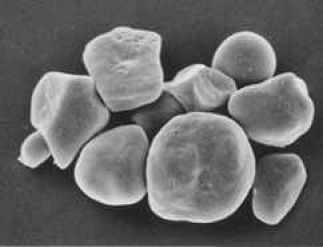
Granules d'amidon de maïs ci-dessus
Micrographie obtenue au microscope à balayage (x3000). (Reproduite avec l'autorisation du Dr. Zoe Ann Holmes, Nutrition and Food Management,Oregon State University,Corvallis, USA).
Complément : Observer
Observer le phénomène de la croix noire
La microscopie en lumière polarisée met en évidence l'arrangement radial des molécules dans le grain d'amidon.
Cette forme du glucose, que l'eau ne peut que très difficilement pénétrer, limite les problèmes de pression osmotique que poserait l'accumulation d'une même quantité de glucose libre dans des organites limités par des membranes.
Complément : Observer
Observer l'ultrastructure de grains d'amidon
Une coupe ultrafine montre les strates concentriques de dépôt d'amidon.
Les proportions relatives d'amylose et d'amylopectine influencent les propriétés physiques de l'amidon.
L'amidon est insoluble dans l'eau froide, et forme un empois (dispersion visqueuse) quand on chauffe le mélange. Cette modification est dûe à l'affaiblissement des liaisons intermoléculaires (principalement hydrogène) avec le chauffage, et se fait en trois étapes. Jusqu'à 70°C environ, les grains d'amidon s'hydratent et gonflent. A ces températures c'est essentiellement l'amylose qui passe en solution. Au refroidissement, cette solution d'amylose forme un gel. Avant la mise au point de l'acrylamide, des gels d'amidon (d'amylose en fait) étaient utilisés pour l'électrophorèse des protéines. Si on continue le chauffage jusqu'à 95°C, l'amylopectine restée dans les grains après diffusion de l'amylose donne une solution très visqueuse qui ne gélifie pas au refroidissement. Si la température est maintenue quelques temps à 95°C, la viscosité de la solution chute brutalement avec la reformation de liens intermoléculaires. De l'eau est expulsée de la masse d'amylopectine. C'est la rétrogradation de l'amidon.
Ces propriétés de l'amidon sont utilisées dans l'industrie alimentaire, dans la préparation d'agents gélifiants (amylose) et épaississants (amylopectine), et aussi dans la fabrication des colles et dans l'industrie papetière. On comprend l'intérêt d'une maîtrise des proportions d'amylose et d'amylopectine dans la qualité des amidons naturels (par la génétique et la sélection des plantes).
L'amidon est fabriqué par tous les végétaux supérieurs et s'accumule en grandes quantités chez la plupart des plantes.
L'amidon transitoire joue un rôle dans le métabolisme photosynthétique du carbone, par lequel il est synthétisé dans les plastes au cours de la photosynthèse, puis dégradé pour fournir de l'énergie et des chaînons carbonés pour le métabolisme en période sombre. L'amidon de réserve est synthétisé dans les amyloplastes des tissus de stockage (graines, tubercules) où il s'accumule pendant une des phases du développement (le développement des graines par exemple), puis est dégradé et utilisé pendant une autre phase (la germination).
Le métabolisme de l'amidon est un chemin important au plan quantitatif. Beaucoup de plantes incorporent la plus grosse partie du carbone nouvellement fixé à partir du CO2 de l'air dans leurs réserves d'amidon.
L'amidon n'est pas l'unique polysaccharide de réserve trouvé chez les végétaux. Quelques plantes, et aussi quelques bactéries, mettent en réserve des polymères de fructose appelés fructanes.
Complément : Les fructanes, quelques exemples illustrent la diversité structurale de ces molécules.
Les fructanes constituent une réserve métabolique chez environ un tiers des espèces végétales.
A côté de l'amidon, les fructanes constituent un autre groupe de polysaccharides de réserve chez les végétaux. C'est le type dominant de réserves dans plusieurs espèces (environ un tiers) du règne végétal. Ils sont naturellement présents en solution dans les vacuoles des cellules, ce qui les distingue de l'amidon qui, lui, est séquestré dans des plastes sous forme cristalline hautement insoluble.
Comme leur nom l'indique, les fructanes sont des polymères de fructose dont le représentant le plus connu est l'inuline, découverte chez Inula helenium. Une particularité cependant est qu'un unique résidu glucopyranosyle est lié à de multiples unités de fructose (jusqu'à 100 000 unités dans certains fructanes bactériens).
On distingue différents groupes de fructanes par les liaisons glycosidiques qui unissent les résidus fructofuranosyles.
les inulines sont des fructanes linéaires avec des liaisons β(2 \(\rightarrow\) 1) entre les unités de fructose. Ce type correspond aux fructanes trouvés chez les dicotylédones et chez quelques monocotylédones.
les lévanes sont aussi des fructanes linéaires mais avec des liaisons β(2 \(\rightarrow\) 6). On les trouve principalement chez des monocotylédones et des bactéries.
les fructanes mixtes (graminanes) sont ramifiés car ils possèdent les deux types de liaisons des fructanes précédents. On les trouve plus spécifiquement chez les graminées, d'où leur nom.
Fabriqués à partir du saccharose dont ils conservent un motif à l'une de leurs extrémités, les fructanes n'ont aucun pouvoir réducteur.
Principaux types de fructanes
Les fructanes diffèrent par la liaison glycosidique unissant les résidus fructofuranosyles. Toutes les molécules de fructanes ont en commun, à une de leurs extrémités, un motif de saccharose (encadré), dont ils dérivent métaboliquement.
Les fructanes protègent les plantes contre le froid et la sécheresse.
Ce n'est sans doute pas un hasard si les plantes à fructanes se trouvent principalement dans les régions à climat froid et/ou sec. Ces molécules pourraient jouer un rôle dans la résistance des plantes aux stress hydrique et thermique. Au contraire de l'amidon, les fructanes sont solubles dans l'eau et osmotiquement actifs (rôle osmoprotecteur contre la déshydratation). En abaissant le point de congélation du liquide vacuolaire, ils ont aussi un rôle cryoprotecteur.
L'amidon fournit plus de la moitié des ressources alimentaires de l'humanité.
A l'échelle mondiale, la plus grande partie de l'amidon alimentaire est fournie par le maïs, mais d'autres sources comme le riz, le blé, la pomme de terre et la patate douce peuvent prédominer selon les régions.

Q uelques produits alimentaires contenant de l'amidon
Le maïs, la pomme de terre et la patate douce sont, avec le riz et le blé (non représentés sur la photo) les principales sources d'amidon alimentaire de l'humanité.
Amylose
L'amylose est un α-D-glucane pratiquement linéaire.
L'amylose est un polymère peu ou pas ramifié d'environ 100 à 1000 résidus de D-glucopyranose (degré de polymérisation moyen DP = 600), enchaînés par des liaisons glucosidiques α(1 \(\rightarrow\) 4).
Complément :
A - Représentation conformationnelle d'un fragment d'amylose
B - Schéma d'une molécule. L'unique extrémité réductrice est figurée par un cercle rouge.
La structure cristalline en double hélice parallèle à enroulement sénestre est celle qui prévaut dans les grains d'amidon. C'est une hélice à 6 résidus par tour, de 2,14 nm de pas. La translation axiale pour un résidu est de 0,35 nm. Il est vraissemblable, mais non vérifié, que des liaisons hydrogène entre groupements -OH de résidus voisins stabilisent la double hélice.
Complément :
A - Un brin isolé - six résidus formant un tour d'hélice sont numérotés -.
B - Hélice double (enroulement sénestre)
C - Hélice double en modèle compact
Dans le cristal d'amylose, l'empaquetage des hélices doubles est de type parallèle (voir Structures de polysaccharides). On connaît plusieurs formes cristallines de l'amylose (A, B et C) qui ne diffèrent que par la densité de l'empaquetage des hélices.
L'amylose est pratiquement insoluble dans l'eau froide. Dans l'eau chaude (entre 50 et 70°C), les molécules se séparent les unes des autres en alternant sur leur longueur des segments désordonnés et des segments hélicoïdaux encore ordonnés. Au refroidissement, ce sont ces segments hélicoïdaux qui, en se réassociant, forment un gel, c'est à dire un réseau moléculaire à mailles très lâches dans lequelles circule le solvant.
L'amylose donne avec l'iode une coloration bleue caractéristique.
L'amylose forme avec l'iode des complexes bleu-noirs très caractéristiques. L'amylopectine ne forme que des complexes violet-rouges instables, et les petits oligosaccharides de moins de 9 résidus ne se colorent pas du tout. Il semble qu'un minimum de 18 résidus soit nécessaire à l'obtention d'une coloration significative.
On sait maintenant qu'en présence d'iode la molécule d'amylose possède une structure différente de la structure native. L'hélice est plus large et ménage un canal central dans lequel les molécules d'iode peuvent s'empiler. L'empilement (et donc l'intensité de la coloration) est d'autant plus efficace que l'hélice est longue, ce qui explique les différences de coloration produites par différents polysaccharides.
Complément :
A - Modèle en squelette de l'hélice d'amylose laissant voir les molécules d'iodes I2 empilées dans la cavité centrale.
B - Cet empilement n'est pas possible dans l'amylose standard (vue axiale), qui ne laisse aucun espace au centre de l'hélice.
C - Dans cette hélice élargie (vue axiale), dont la cavité centrale s'ajuste exactement aux dimensions de l'iode, cet empilement devient possible...
Amylopectine
L'amylopectine est une version ramifiée de l'amylose.
Comme l'amylose, l'amylopectine est un α(1 \(\rightarrow\) 4)-glucane, mais des ramifications sont attachées via des liaisons glucosidiques α(1\(\rightarrow\) 6) à des chaînes linéaires de résidus glucopyranosyles liés en α(1\(\rightarrow\) 4). Ces ramifications interviennent en moyenne tous les 20-25 résidus sur une chaîne principale, et elles contiennent environ 15-30 résidus liés en α(1\(\rightarrow\) 4). Certaines ramifications sont elles-mêmes ramifiées. Les molécules d'amylopectine isolées des cellules végétales contiennent en moyenne 2000 unités de glucose (DP = 2000) mais certaines peuvent en contenir jusqu'à 6000. La molécule dans son ensemble présente un aspect arborescent.
Complément :
A - Représentation conformationnelle
B - Schéma d'une molécule. L'unique extrémité réductrice est figurée par un cercle rouge. La chaîne principale est représentée en vert et les ramifications en orange.
La structure tridimensionnelle de l'amylopectine dans les grains d'amidon n'est pas encore connue.
Des modélisations réalisées sur ordinateur suggèrent l'existence d'une structure hélicoïdale à enroulement sénestre, voisine de celle de l'amylose et typique des α(1 \(\rightarrow\) 4)-glucanes. Conformément à ce que l'on sait de la liberté conformationnelle permise par les liaisons glycosidiques 1\(\rightarrow\) 6 (voir Diversité de formes des oligosaccharides), les points de branchement ne semblent pas s'opposer à l'enroulement hélicoïdal des chaînes. La coloration rouge-violet de l'amylopectine par l'iode (en contraste avec la coloration bleu-noir de l'amylose) confirme l'existence de segments d'hélices beaucoup plus courts.
Complément :
La structure modélisée correspond à un point de branchement entre une chaîne de 6 résidus (1 à 6) et une chaîne de 4 résidus (1' à 4'). En tenant compte des règles de calcul en usage pour la modélisation de telles structures, l'ordinateur fait clairement apparaître un enroulement hélicoïdal sénestre aussi bien pour la chaîne de 4 résidus (point de vue A) que pour la chaîne de 6 résidus (point de vue B)
Dans le grain d'amidon, l'amylopectine forme des tablettes noyées dans la masse d'amylose. Dans chaque tablette, les molécules d'amylopectine sont parallèles les unes aux autres, toutes orientées de la même manière, avec leurs extrémités réductrices dirigées vers le centre du grain. Dans chaque tablette, il est probable que des petits segments d'hélices doubles se forment entre molécules voisines.
Complément :
Dans chaque tablette, les molécules d'amylopectine sont empaquetées parallèlement les unes aux autres, leurs extrémités réductrices (cercles rouges) dirigés vers le centre du grain d'amidon. Des hélices doubles pourraient se former entre segments voisins. .
Hydrolyse enzymatique
L'hydrolyse de l'amidon par les acides dilués se fait lentement. Elle libère dans un premier temps de grands fragments appelés dextrines qui ne forment plus d'empois au chauffage. Le raccourcissement progressif des dextrines, jusqu'à l'obtention de glucose libre, peut être suivi par la coloration à l'iode qui passe successivement du violet au rouge, puis disparaît quand les fragments deviennent trop petits (moins de 10 résidus).
Amylose et amylopectine sont hydrolysés par des amylases.
Les végétaux, les animaux et les micro-organismes possèdent une batterie d'enzymes (glucosyl hydrolases) capables d'hydrolyser l'amidon, et de libérer le glucose qu'il contient.
L'α-amylase, qui existe aussi bien chez les animaux que chez les végétaux, et aussi les bactéries, est une endo-glucosidase qui catalyse l'hydrolyse aléatoire des liaisons glucosidiques α(1\(\rightarrow\) 4) de l'amylose et de l'amylopectine. Les produits d'hydrolyse sont des oligoholosides de 6 à 7 résidus en moyenne et un disaccharide, l'α-D-glucosyl-(1\(\rightarrow\) 4)-D-glucose, ou maltose.
Structure du maltose
(Représentation conformationnelle de Reeves). Le maltose est un α-D-glucosyl-(1 \(\rightarrow\) 4) -D-glucose issu de l'hydrolyse enzymatique de l'amidon. C'est un disaccharide réducteur dont la forme anomérique majoritaire est β, comme pour le glucose libre.
Structure de l'isomaltose
(Représentation conformationnelle de Reeves). L'isomaltose est un α-D-glucosyl-(1 \(\rightarrow\) 6)-D-glucose issu de l'hydrolyse enzymatique de l'amidon. C'est le motif constituant les points de branchement dans l'amylopectine et le glycogène.
Une autre hydrolase, la β-amylase, est trouvée dans les graines et les tubercules de certaines plantes supérieures, ainsi que chez des bactéries. La β-amylase est une exo-glucosidase qui catalyse l'hydrolyse de la liaison glucosidique α(1 \(\rightarrow\) 4) à partir des extrémités libres, non-réductrices, de l'amylopectine, et libère séquentiellement du maltose. Noter que le préfixe β- de cette amylase n'a rien à voir avec la configuration de la liaison hydrolysée. C'est simplement la préférence anomérique du maltose libéré qui est rappelée.
Les liaisons α(1 \(\rightarrow\) 6) de l'amylopectine n'étant pas reconnues par la β-amylase, l'hydrolyse s'arrète quand l'enzyme atteint les points de branchement. La molécule d'amylopectine partiellement hydrolysée est appelée dextrine limite. Elle ne peut être ultérieurement dégradée qu'après l'action d'enzymes spécifiques des liaisons glucosidiques α(1 \(\rightarrow\) 6), les enzymes débranchantes ou déramifiantes.
Les enzymes déramifiantes les mieux connues sont d'origine végétale ou microbienne (isomaltase, pullulanase). In vitro, elles agissent aussi bien sur l'amylopectine que sur le glycogène. L'isomaltase humaine hydrolyse l'isomaltose (α-D-glucosyl-[1 \(\rightarrow\) 6]-D-glucose) et le maltose (α-D-glucosyl-[1 \(\rightarrow\)4]-D-glucose), ainsi que de petites dextrines.
Complément : Observer
Observer un grain d'amidon partiellement hydrolysé
Au microscope à balayage, l'hydrolyse différentielle de l'amylose et de l'amylopectine fait particulièrement bien ressortir les strates moléculaires.
L'amidon peu hydrolysé peut encore être fragmenté par des cyclodextrines glycosyltransférases, synthétisées par quelques bactéries (Bacillus macerans, B. subtilis etc...). Cette dégradation enzymatique produit des anneaux de 6 à 8 résidus de glucose, les cyclodextrines.
Complément : Les cyclodextrines
Les cyclodextrines sont produites par dégradation enzymatique de l'amidon.
Les cyclodextrines sont des oligosaccharides cycliques produits industriellement. Ce ne sont donc pas des molécules naturellement présentes dans les cellules, mais leur intérêt industriel mérite d'en faire mention ici. Les cyclodextrines sont composées de 6 et plus résidus D-glucosyle. Elles sont classées par le nombre de résidus qu'elles contiennent : les α-cyclodextrines en contiennent 6 (cyclomaltohexaose), les β-cyclodextrines 7 (cyclomaltoheptaose), et les γ-cyclodextrines 8 (cyclomaltooctaose). La structure moléculaire d'une β-cyclodextrine est illustrée ci-dessous.
Modèle moléculaire de β-cyclodextrine (cyclomaltoheptaose)
Vue de face (à gauche), la molécule ressemble à un anneau, formé de 7 rédisus α-D-glucosyle. Vue de profil (à droite, encadré), la molécule est nettement de forme tronc-cônique, avec tous les alcools primaires (CH2OH) situés sur la petite section, et tous les alcools secondaires (-OH) sur la grande section.
Les enzymes reponsables de la formation des cyclodextrines sont les cyclodextrine-glycosyltransférases. Ces enzymes sont produites par des bactéries comme Bacillus circulans qui les excrètent dans le milieu extracellulaire quand on fournit de l'amidon (voir Amidon) comme unique source de carbone. Elles sont optimisées par génie génétique pour favoriser la formation d'une classe particulière de cyclodextrine. De plus, des agents de précipitation sélectifs sont utilisés pour isoler les cyclodextrines α, β et γ. Plusieurs milliers de tonnes de cyclodextrines sont produites de cette manière dans le monde chaque année.
L'intérêt des cyclodextrines réside dans leur aptitude à inclure diverses molécules.
Les cyclodextrines sont des molécules chirales, solubles dans l'eau, biodégradables, accessibles aux modifications chimiques, et de plus capables d'inclure d'autres molécules dans leur cavité centrale relativement hydrophobe. Un grand nombre de substances actives comme des drogues anti-cancéreuses ou anti-inflammatoires, des hormones, des colorants, des arômes, peuvent êtres incluses dans les cyclodextrines, pour augmenter leur solubilité dans l'eau et pour contrôler leur libération dans les milieux. Les médicaments insolubles dans les fluides biologiques deviennent plus solubles, plus stables et mieux protégés. De telles associations sont appelées complexes d'inclusion. On peut aussi complexer des polymères de grande taille avec de nombreux anneaux de cyclodextrine, et ainsi altérer leurs propriétés physico-chimiques.
Cette représentation respecte les proportions des atomes et montre bien la forme de la molécule. La molécule est vue par la grande section du tronc de cône, porteuse des alcools primaires (en rouge et gris clair). La taille de la cavité centrale d'une β-cyclodextrine est suffisante pour inclure une molécule de la taille d'un noyau naphtalène.
Les cyclodextrines s'emploient dans les industries alimentaire, pharmaceutique, cosmétique.
Les amylases sont très utilisées dans l'industrie.
Les applications majeures des amylases dans l'industrie alimentaire concernent la brasserie et la boulangerie, qui nécessitent une dégradation préalable de l'amidon contenu dans les grains en petits fragments utilisables (on dit fermentescibles) par les levures.

Utilisation des amylases en boulangerie (ci dessus)
Au cours du pétrissage et de la levée du pain, les amylases naturellement présentes dans les grains de blé sont activées par la réhydratation de la farine. Le glucose libéré par leur action est utilisé par les levures, et produit de l'éthanol et du CO2 par fermentation alcoolique. Au cours de la cuisson du pain, l'éthanol s'évapore, et le gaz piégé forme les bulles de la mie. Par ailleurs, la réaction du glucose avec des acides aminés issus de la dégradation des protéines, donne naissance à des composés aromatiques responsables du goût caractéristique du pain cuit.
Les amylases sont également utilisée en confiserie dans la préparation de sirops à partir d'amidon. Le sirop de maïs contient plus de 70% de sucre sous forme de glucose, de maltose et d'oligoholosides de diverses longueurs. L'isomérisation enzymatique (voir Isomérisation) d'une partie du glucose en fructose crée un mélange dont la texture et le pouvoir sucrant s'approchent de ceux du sucre inverti (voir Saccharose et tréhalose).