Structures secondaires _ Cambium
Chez les Gymnospermes et chez les plantes à fleurs (ANGIOSPERMES Dicotylédones) se développent des formations secondaires grâce aux méristèmes secondaires : les CAMBIUMS.
Le développement d'une Dicotylédone, par exemple, se fait donc en deux temps :
Tout d'abord, les méristèmes primaires fonctionnent seuls mettant en place des formations primaires,
puis, après un laps de temps plus ou moins important suivant les espèces, aux formations primaires viennent s'ajouter des formations secondaires produites par les cambiums.
A l'origine, un cambium se présente comme un cylindre de cellules indifférenciées s'étendant sur tout ou partie de l'organe où il se trouve.
Ce cylindre est généralement constitué d'une zone cambiale, regroupant plusieurs couches de cellules.
I _ Les deux cambiums
Evolutivement, deux types de cambium ont émergé chez les Dicotylédones :
Le cambium xylémo-phloémien.
Encore appelé cambium vasculaire ou cambium, c'est le plus important pour le métabolisme d'une plante à formations secondaires. Apparaissant au sein des éléments conducteurs du cylindre central, il produit, souvent en abondance, de nouveaux éléments conducteurs : xylème et phloème secondaire.
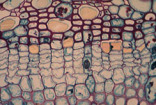

Fig. 01 : Photographie et détail d'une zone cambiale vasculaire
Le cambium subéro-phellodermique ou phellogène.
Moins courant que le précédent, n'intervenant que tardivement dans le cas des arbres, il se forme dans l'écorce. La plante lui doit deux tissus secondaires :
Un tissu de revêtement, le suber (liège), qui remplacera le rhizoderme exfolié dans une racine âgée et qui donnera un tissu protecteur complexe quand la tige vieillira;
Le phellogène produit également - pas toujours - un tissu parenchymateux à fonction d'accumulation de réserves essentiellement : le phelloderme.
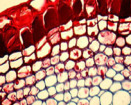
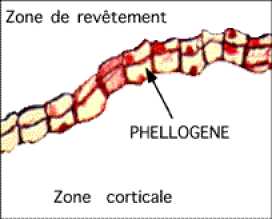
Fig. 02 : Photographie et détail d'un phellogène
Les botanistes, depuis fort longtemps, se sont intéressé au cambium vasculaire à cause de son importance économique : industries du bois et du papier.
De nombreux résultats significatifs ont vu le jour.
Plusieurs questions restent encore sans réponse malgré tout. Les domaines d'étude portent sur la structure de ce cambium et sur son mode de fonctionnement.
II _ Structure du cambium vasculaire
Y a-t-il relation entre procambium et cambium ?
C'est la première question qui se pose à propos de ces deux méristèmes. Le cambium apparaît au sein des tissus procambiaux plus ou moins tardivement. Il ne peut se développer que de deux façons : soit à partir de certaines cellules de ce procambium, soit à partir de cellules déjà déterminées en attente dans le procambium.
Au cours du développement, il semble y avoir continuité entre les fonctionnements des deux méristèmes, sans que la question soit clairement tranchée aujourd'hui.
Anatomie du cambium.
Les travaux de Bailey [(1919-1930) et des travaux plus récents (1954)]* ont permis de la préciser chez une Gymnosperme : Pinus strobus. Contrairement à ce que l'on peut observer dans un méristème primaire, deux sortes de cellules initiales y figurent : les initiales fusiformes et les initiales radiales. Les premières donneront naissance à des éléments conducteurs qui s'orientent longitudinalement, dans le sens de l'axe de la plante. Les secondes seront à l'origine des rayons xylémo-phloémiens conduisant les sèves du centre à la périphérie (et vice versa) de l'organe.
["Les premières" donneront naissance, chez les Gymnospermes : aux cellules conductrices (trachéïdes) et éventuellement aux canaux sécréteurs de résine; chez les Angiospermes: aux cellules conductrices (vaisseaux), aux cellules associées (ex. cellules compagnes), aux fibres, aux cellules de parenchyme vertical.]*
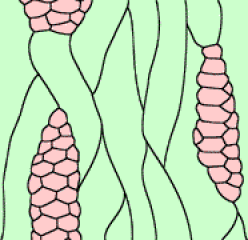
Productions cambiales.
On sait qu'il s'agit de xylème et de phloème dits secondaires.
Les initiales, par division, conduisent à la production de xylème de la périphérie vers le centre (différenciation centripète) et de phloème de la moelle vers le péricycle (différenciation centrifuge).
Les éléments secondaires étant alignés en files radiales (d'où le nom ancien de liber=livre), il a été longtemps admis que les initiales ne pouvaient se diviser que sur le mode péricline. En observant des mitoses anticlines, Nageli [(1858), confirmé par Bailey]* a résolu un problème ayant longtemps divisé les botanistes.
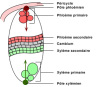
III _ Répartition et forme du cambium vasculaire
Cambium intra et inter-fasciculaire.
Chez certaines plantes (Tilia), d'emblée, le cambium xylémo-phloémien forme un cylindre continu se déployant entre phloème primaire et xylème primaire. Il y est à la fois intra et inter-fasciculaire.
Chez d'autres (Medicago) il reste intra-fasciculaire.
Assise ou zone cambiale ?
Il s'agit également d'une question ayant longtemps prêté à controverse.
N'y a-t-il qu'une seule couche de cellules ayant valeur de cellules cambiales, où s'agit-il plutôt d'une zone regroupant plusieurs assises de cellules ayant toutes les caractéristiques du cambium ?
Le consensus des botanistes semble se faire aujourd'hui sur la seconde idée.
Organisation cambiale.
Trois zones sont à distinguer :
La zone des initiales cambiales. Plusieurs couches de cellules à divisions anticlines et périclines la constituent.
Une zone de cellules-mères du phloème et une autre de cellules-mères du xylème encadrent la zone des initiales. Les divisions anticlines s'y raréfient jusqu'à disparaître tandis que les divisions périclines y sont plus nombreuses.
Tout à fait à l'extérieur, enfin, phloème et xylème secondaires sont plus ou moins abondants dans leur forme mature.
Zone de différenciation du phloème
CAMBIUM
Cellules-mères phloèmiennes
Initiales cambiales
Cellules-mères xylèmiennes
Zone de différenciation du xylème