Proembryon
Post-fécondation
Chez Arabidopsis thaliana et chez Capsella bursa-pastoris, plantes communes appartenant à la famille des Brassicacées, la structure de l'ovule (Fig. 01) est nettement polarisée. On peut y définir un axe de symétrie bilatérale passant par le micropyle (M) à un pôle, et par la chalaze (C) à l'autre pôle.
Le gynécée de chaque fleur renferme généralement plusieurs de ces ovules qui, par le jeu de la pollénisation, seront fécondés ou non. Celà dépendra du nombre de grains de pollen arrivant sur le stigmate, de leur degré de maturité, de l'accessibilité des ovules eux-mêmes, etc...
C'est ce gynécée qui va se transformer en fruit après anthèse. C'est donc à l'intérieur de ce fruit (une silicule chez Capsella bursa-pastoris) que se retrouveront les graines (Fig. 02), dans lesquelles il conviendra d'observer les embryons.
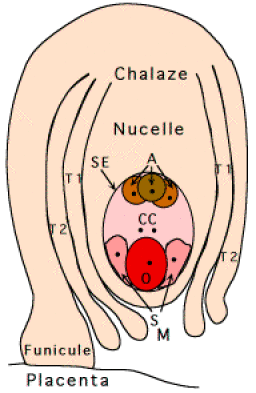
T1 = Tégument externe (exine)
T2 = Tégument interne (intine)
CC = Cellule Centrale
SE = Sac Embryonnaire
A = Cellules Antipodiales
O = Oosphère
S = Synergides
M = Micropyle
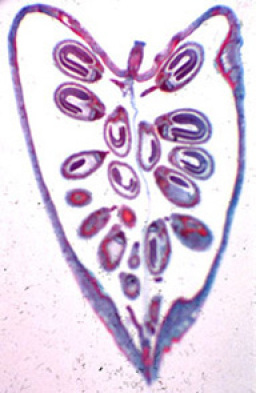
Le fruit de Capsella bursa-pastoris est une silicule. Vu en coupe sur ce cliché, il contient un certain nombre de graines contenant des embryons à divers stades de maturité.
L'embryogenèse débute au moment où le zygote principal entre en division par des mitoses successives.
La première mitose clive le zygote en deux cellules-filles différentes : A= Apicale; B=Basale.
La cellule A du côté chalazial est plus petite, la cellule B du côté micropylaire le long de l'axe de symétrie ovulaire est plus volumineuse. Avant qu'une nouvelle division n'intervienne, la surface de A directement au contact du nucelle se subérifie devenant ainsi imperméable. Un sens bien précis de transfert des substances nutritives s'établit alors entre nucelle, cellule A et cellule B. Aucun élément ne pourra pénétrer en A sans passer nécessairement par B, qui exerce ainsi un rôle de filtre pour les substances nutritives destinées à l'embryon.
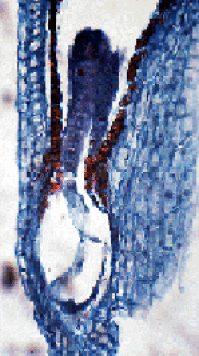
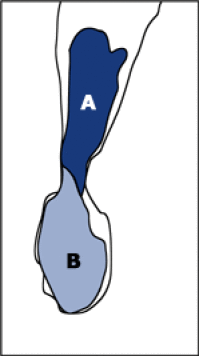
Fig. 03 : Photographie et schéma d'un embryon de Capsella bursa-pastoris après la première mitose du zygote.
La seconde mitose divise A et B en même temps, produisant 4 cellules-filles.
Ces deux nouveaux plans de division étant anticlines l'un à l'autre, les nouvelles cellules se disposent disymétriquement.
Troisième série de mitoses.
Les 4 cellules-filles provenant de B forment un filament cellulaire, les divisions y étant toutes périclines. L'organe ainsi constitué est le suspenseur. En A, par contre, les divisions anticlines disposent les 4 cellules-filles côte à côte. Il s'agit de l'embryon proprement dit, appelé pro-embryon, à ce stade précoce de son développement.
Un peu plus tard, le suspenseur deviendra un filament d'une dizaine de cellules. Il sera chargé de nourrir l'embryon en lui transférant les substances nutritives qu'il puise dans le nucelle.
emb-globulaire
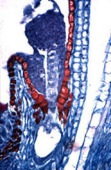
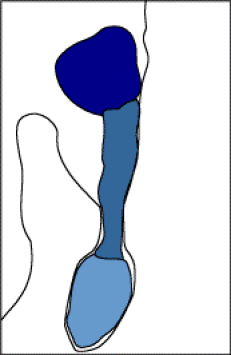
Fig. 04 : Photographie et schéma d'un embryon de Capsella bursa-pastoris au stade globulaire.
Le pro-embryon se présentera sous la forme d'une petite sphérule depuis 16 cellules. C'est le stade globulaire où apparaît une certaine différenciation :
Une première assise de cellules superficielles formera à terme les tissus de revêtement de la plante.
Au cœur du pro-embryon, quelques cellules constituent le procambium, ébauche du cylindre central.
La zone intermédiaire donnera naissance à l'écorce.
Transition
Elle est caractérisée par la mise en place de deux éléments importants de la plante à naître :
L'allongement du pro-embryon, d'une part, enclenche la mise en place de l'axe du végétal.
Deux expansions latérales, d'autre part, produisent les deux cotylédons.