Division
L'une des caractéristiques majeures des êtres vivants est la reproduction. L'objectif de la Vie est de se "répandre sur Terre", "d'occuper les espaces libres" dira l'écologue.
L'instrument forgé par les plantes (comme par les animaux) pour atteindre cet objectif est toujours le même : augmenter le nombre de cellules, ou "unités de vie", en les dotant du pouvoir de se diviser.
Au cours du Temps, de plus en plus raffiné et efficace, divers modes de division ont émergé.
Ce sont les bactéries (toutes les cellules procaryotes plus précisément) qui sont apparues les premières dans la biosphère, mettant en oeuvre le mode de division le plus simple (Fig. 01).
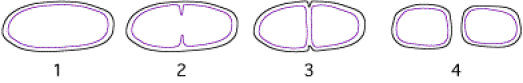
Après un temps de vie relativement court (une trentaine de minutes pour une bactérie), une constriction apparaît dans la paroi (2). Gagnant du terrain vers l'intérieur de la cellule, cette constriction, comme un diaphragme d'appareil photographique qui se ferme, sépare les constituants de la cellule d'origine en deux lots (3). Ce mode de division s'appelle la scissiparité.
Chez les Eucaryotes, la division cellulaire est plus complexe. Elle assure une parfaite parité génétique entre les deux cellules filles produites. C'est le mécanisme connu chez les plantes et chez les animaux sous le terme de MITOSE.
L'organisme d'un végétal supérieur est constitué le plus souvent d'un grand nombre de cellules assemblées en organes divers. Toutes ces cellules n'ont pas le même comportement au cours du développement en ce qui concerne le processus de division.
Certaines, après leur formation, ne cesseront pas de se diviser plus ou moins régulièrement pendant toute la durée de vie de l'organisme qui les abrite. C'est le cas de certaines cellules des méristèmes végétaux,
D'autres, au contraire, ne se diviseront jamais après leur formation dans l'embryon. Elles se différencient très vite et ne changeront pas d'état jusqu'à la disparition de la plante,
La majorité des cellules du végétal, cependant, après s'être divisées un certain nombre de fois, perdent leur pouvoir mitotique et ne donneront plus de cellules-filles.
Chacune des cellules, pour sa part, voit se succéder de nombreux événements entre la mitose qui lui donne naissance et celle qui la scindera en deux cellules-filles.
La période de temps qui s'écoule entre ces deux mitoses est le cycle cellulaire. Il se termine par la mise en place des éléments préparatifs de la future mitose,
Le positionnement de la future paroi prend une importance capitale chez les végétaux. Les mécanismes qui le contrôlent seront décrits,
La division du noyau (cytodiérèse) peut alors se dérouler. Cette phase est étudiée au chapitre 3 consacré à la cellule végétale,
La construction de la nouvelle paroi (cytokinèse), enfin, est réalisée, séparant définitivement les deux cellules-filles.
Remarque : Cytodiérèse et cytokinèse sont réunies sous le terme de mitose.
I _ Le cycle cellulaire
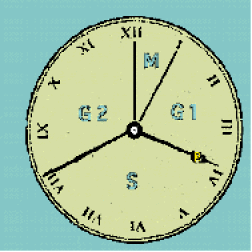
Il y a peu, l'idée générale était que, entre deux mitoses successives, la cellule passait par une phase de repos nommée interphase. Le développement des connaissances permet aujourd'hui d'élaborer un modèle plus complexe. L'interphase est en réalité un temps de préparation active de la mitose. La nature des évènements qui y prennent place incite à découper arbitrairement l'interphase en trois parties :
Phase G1 _ Après la mitose, la taille de chaque cellule-fille ayant été fortement réduite, G1 est caractérisée par une croissance cellulaire active chargée d'amener chaque cellule-fille à la même taille que la cellule-mère originelle. Le nombre et la taille des organites sont augmentés. Dans le noyau, la chromatine est en voie de décondensation.
Phase S _ On la fait débuter au moment où la cellule commence à répliquer son matériel génétique totalement décondensé. La fin de la réplication sert également à repérer le moment où S se termine. A la limite entre G1 et S se place un point particulier du cycle : le point de restriction R.
Il désigne un seuil au delà duquel la division devient irréversible : quelles que soient les conditions externes ou internes à la cellule, la division ira à son terme si R a été franchi.
On constate alors que R joue un rôle régulateur qui permettra à une cellule d'allonger son cycle en retardant tout simplement le franchissement du point de restriction.
Phase G2 _ La réplication de son ADN étant achevée, une dernière étape consistera pour la cellule à préparer une nouvelle mitose, notamment en synthétisant les diverses molécules qui lui seront nécessaires à cet effet et en condensant sa chromatine jusqu'au stade de chromosome individualisé visible en microscopie.
II _ Le positionnement de la paroi
L'une des caractéristiques remarquable du végétal est son manque de mobilité. A tous les niveaux de son organisation, chaque élément meurt généralement à l'endroit précis ou il est apparu : l'individu sur la portion de sol où il s'est enraciné, la branche secondaire sur la branche maîtresse qui la porte, la fleur sur son pédoncule.
Cette absence de mouvement se remarque même au niveau cellulaire car, en effet, il n'y a pas de migration des cellules chez une plante. Ceci est dû à la présence de la paroi pecto-cellulosique qui sépare et réunit, en même temps, deux cellules contiguës.
Ainsi, au terme d'une division cellulaire chez une plante, les deux cellules-filles néoformées resteront unies l'une à l'autre par la nouvelle paroi qui s'interpose entre elles.
Ceci donne une grande importance au choix de la position où se formera cette paroi dans le volume de la cellule-mère.
En apparence, le cytosquelette semble jouer le rôle principal dans ce choix.
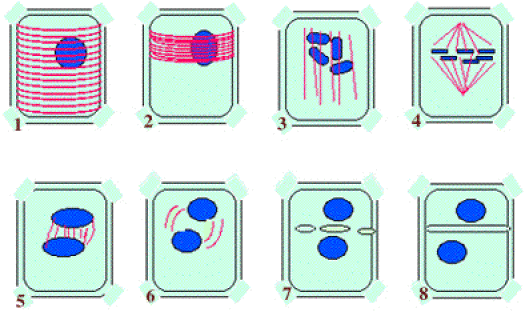
Dans une cellule encore en interphase, les microtubules, à l'intérieur de la cellule, sont uniformément répartis à la périphérie, sous le plasmalemme ;
Le début de la prophase mitotique est indiqué par un événement significatif au niveau des microtubules périphériques : ils se concentrent suivant l'un des diamètres de la cellule qui va se diviser.
La figure qu'ils constituent ainsi a reçu le nom de "bande préprophasique".
Entraîné par les filaments d'actine qui le relient au plasmalemme, le noyau se positionne au centre de la bande préprophasique.
Le noyau de la cellule parcourt la prophase de la mitose. La condensation des divers chromosomes qui le constituent touche à sa fin, et les chromosomes deviennent distincts les uns des autres.
Pendant ce temps, les microtubules se disposent longitudinalement dans la cellule. Ils formeront bientôt, après réorganisation, le "fuseau achromatique".
Ce fuseau atteint sa maturité à la métaphase. Les chromosomes, au maximum de leur condensation et de leur visibilité se disposent en "plaque équatoriale" dans la zone centrale du fuseau achromatique.
Les deux noyaux qui feront vivre les deux cellules-filles sont reconstitués. Les microtubules, du fuseau désorganisé, regagneront graduellement la périphérie des cellules nouvellement formées.
Les deux noyaux qui feront vivre les deux cellules-filles sont reconstitués. Les microtubules, du fuseau désorganisé, regagneront graduellement la périphérie des cellules nouvellement formées.
Après la cytodiérèse, la cytokinèse est enclenchée et la nouvelle paroi va, par le biais du phragmoplaste, séparer les deux cellules issues de la mitose.
Après la cytodiérèse, la cytokinèse est enclenchée et la nouvelle paroi va, par le biais du phragmoplaste, séparer les deux cellules issues de la mitose.
Ainsi, dans un organe végétal en développement, suivant que les mitoses disposent les parois néoformées de telle ou telle façon, la morphologie de l'organe variera :
Si les nouvelles parois sont disposées parallèlement les unes aux autres, les mitoses sont dites "périclines" (voir ci-dessous).


Si les nouvelles parois sont disposées perpendiculairement les unes aux autres, les mitoses sont dites "anticlines" (Fig. 05).

Si les nouvelles parois sont disposées aléatoirement les unes aux autres, la forme de l'organe sera elle-même "aléatoire" (Fig. 06).
III _ Cytodiérèse (les différentes phases de la mitose)
Les clichés ont été pris sur une préparation de racine de jacinthe. Les racines sont fixées, inclues dans la paraffine puis sectionnées longitudinalement. Les coupes sont ensuite traitées par le test de Feulgen qui colore spécifiquement l'ADN en rouge. Afin de mieux visualiser la structure cellulaire, une coloration d'appoint au vert lumière est ajoutée. Le cytoplasme (fibres du fuseau incluses) et les nucléoles sont alors colorés en vert. La paroi et les vacuoles ne sont pas colorées.

Les noyaux apparaissent en rouge. Malgré le faible grossissement, on observe quelques figures de mitose.
A partir de cette coupe, les différents stades de la mitose ont été sélectionnés puis photographiés à fort grossissement :
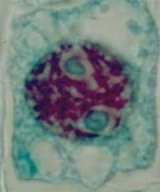
Le noyau est délimité par l'enveloppe nucléaire. Il contient de l'ADN sous forme de chromatine disposée sans régularité apparente.
On observe deux nucléoles. Le cytoplasme de cette cellule méristématique est légèrement vacuolisé
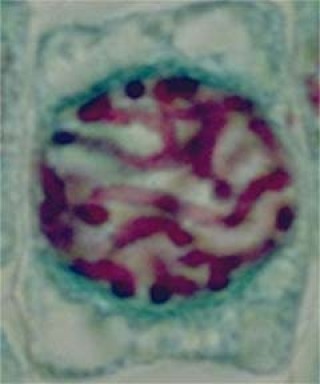
Les chromosomes s'individualisent à partir de la chromatine. Les nucléoles et l'enveloppe nucléaire disparaissent
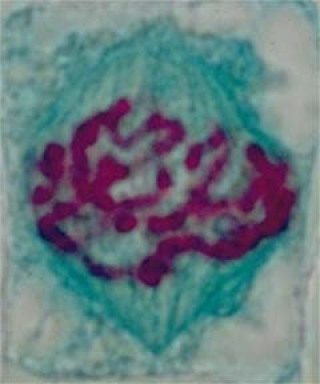
Le fuseau formé de fibrilles (microtubules) se forme autour des chromosomes qui sont regroupés au centre. Certaines fibres fusoriales s'accrochent aux centromères des chromosomes.
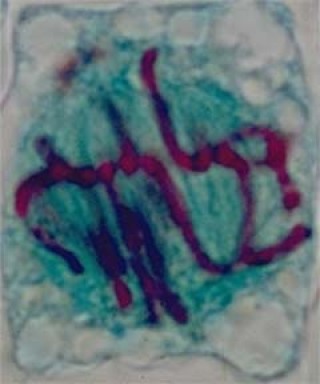
Les chromosomes, tirés par les fibres fusoriales, se regroupent à l'équateur du fuseau (plaque équatoriale). Rien d'autre ne se produit jusqu'à ce que tous les chromosomes soient situés à l'équateur.

Brutalement, tous les chromosomes se clivent au niveau de leur centromère. Chaque "demi-chromosome" ou chromatide est tiré vers un pôle du fuseau par les fibres fusoriales attachées aux centromères.
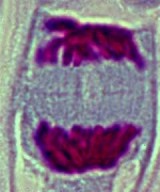
Les chromatides encore bien individualisés se regroupent aux deux pôles du fuseau.
Elles forment deux lots de chromosomes parfaitement identiques. Le fuseau est toujours visible.
Au niveau de l'équateur, des vésicules se rassemblent au centre et forment une plaque (le phragmoplaste).
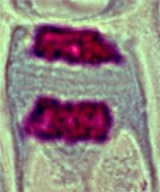
La plaque cellulaire (phragmoplaste) s'étend vers la périphérie. Ce phragmoplaste, constitué principalement de pectines, se relie à la paroi cellulaire préexistante et sépare complétement la cellule en deux. Les chromosomes perdent leur individualité morphologique
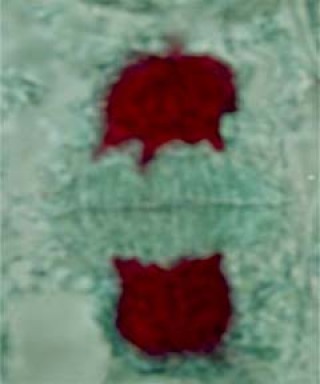
Chaque lot de chromosome va s'entourer d'une nouvelle enveloppe nucléaire et reformer un noyau interphasique.
Ces clichés ont été obtenus à partir de préparations réalisées au service du SNV1 (D.Robert)
IV _ Construction de la paroi : le phragmoplaste
La localisation de la nouvelle paroi étant fixée, la cellule mère en commence la construction :
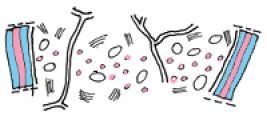
1. Les premiers évènements de cette construction consistent simplement en une accumulation significative, au niveau du diamètre choisi par la cellule, de certains organites : mitochondries productrices d'énergie et dictyosomes chargés des synthèses ;
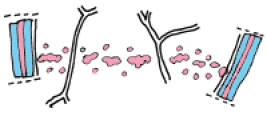
2. Rapidement, le golgi bourgeonne de nombreuses vésicules gorgées de pectine ;
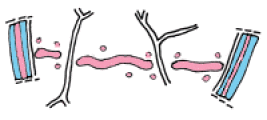
3. L'étape qui suit est caractérisée par la confluence progressive de ces vésicules golgiennes jusqu'à ne former qu'un seul ensemble, une sorte de "galette" de pectine limitée extérieurement par une membrane phospho-lipidique. De place en place, des travées de reticulum endoplasmique ménagent des interruptions ponctuelles dans cette lamelle pectique. Elles formeront à terme les plasmodesmes. A ce stade, déjà, la séparation de la cellule-mère en deux cellules-filles est matérialisée sous la forme de cette lamelle pectique encore appelée lamelle mitoyenne.
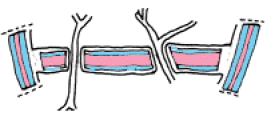
4. Il ne reste plus à chaque cellule-fille qu'à déposer les diverses couches pecto-cellulosiques qui constitueront la paroi nouvelle.