Structures _ Méristèmes primaires
La variabilité des structures méristématiques étant très grande, il est malheureusement impossible de toutes les représenter par un modèle unique. Des différences entre elles se manifestent à tous les niveaux dans l'étude de la plante :
Entre méristèmes primaires et secondaires,
Entre méristèmes caulinaires, radiculaires et foliaires,
Entre méristèmes dans un même organe (mais pour des espèces différentes),
Entre méristèmes jeunes et âgés, également.
L'état indifférencié dans lequel se trouvent en principe toutes les cellules méristématiques d'un organisme, compliquera encore l'analyse des structures : comparer deux masses de cellules indifférenciées est le plus souvent peu parlant, obligeant l'observateur à une analyse très fine de ces tissus.
Seuls quelques éléments de réponse sont actuellement disponibles. Ils concernent les méristèmes principaux d'un végétal.
Cellule et mitose
Ce comportement des cellules végétales amène d'autres réflexions sur le pouvoir mitotique.
Certaines cellules méristématiques restent dans les limites du méristème jusqu'à leur mort. On leur donne le nom d'"initiales".
Généralement, une initiale ne se différencie jamais. Sa seule fonction est de produire des cellules-filles à un rythme plus ou moins soutenu, à des intervalles de temps plus ou moins longs.
On nomme "cellules dérivées des initiales", les cellules-filles auxquelles elles ont donné naissance et qui sont restées dans les limites du méristème.
Hors du méristème, les cellules rencontrées sont diverses.
Certaines continuent à se diviser plusieurs fois avant que les mitoses ne cessent et que la différenciation intervienne, d'autres se différencient directement en tel ou tel type cellulaire.
Une cellule méristématique peut être déterminée dès sa naissance. Quel que soit le nombre de mitoses qu'elle subit préalablement, elle est déjà programmée pour devenir un type cellulaire précis.
Dans ce cas, les divisions successives qu'elle subit produisent des cellules-mères du même type.
D'autres, dites "indéterminées", se divisent plusieurs fois sans que leur devenir soit fixé.
Remarque : Les mécanismes moléculaires fixant la détermination d'une cellule sont encore mal connus.
Méristèmes caulinaires
Une coupe anatomique longitudinale effectuée au niveau de l'apex permet d'étudier la structure du méristème. Suivant le groupe de plantes auquel on s'adresse la figure anatomique obtenue ne sera pas la même.
Structure apicale de Coleus sp.
L'apex montre une organisation repérable des plus simples :
Dans la zone superficielle, la majorité des plans de division à la mitose sont parallèles les uns aux autres et perpendiculaires à la surface de l'organe. Deux plans de division parallèles sont dits "périclines". [Ils sont "anticlines" quand ils font entre eux un angle de 90°]. La zone superficielle de l'apex aura donc tendance à s'organiser en plusieurs couches. On distinguera ainsi, suivant la nomenclature proposée par Satina & al. (1940) les couches L1, L2, L3. Cet ensemble de couches cellulaires aura tendance à augmenter la surface de l'apex. On lui donne le nom de tunica.
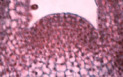

Fig. 02 : Photographie et schématisation d'un apex caulinaire de Coleus sp.
Sous la tunica, les plans de division se disposent aléatoirement dans toutes les directions de l'espace.
La structure obtenue formera une sphérule massive de cellules plutôt qu'un ensemble de couches. C'est le corpus qui augmentera quant à lui le volume de l'apex.
En position sub-apicale à la périphérie du méristème, l'ébauche d'une feuille (ou de plusieurs) est visible. Ce sont des primordiums foliaires.
Sous le corpus, enfin, dans la partie centrale de l'axe en formation, les cellules fortement vacuolisées constituent un méristème médullaire, moelle embryonnaire du futur cylindre central.
La différenciation en Tunica / Corpus semble la plus courante chez les plantes à fleurs (Angiospermes). Chez les Gymnospermes les choses se présentent un peu différement.
Structure apicale dans un apex de Ginkgo biloba.
Au coeur de l'apex s'observe une zone centrale de cellules-mères. Une couche superficielle, à plans de divisions anticlines prédominants la recouvre. A la périphérie de la zone centrale, un anneau de petites cellules à divisions périclines constitue une zone périphérique sub-superficielle.
Sous la zone centrale, un cordon méristématique à parois perpendiculaires à l'axe de l'organisme donnera naissance à une moelle.

cs = couche superficielle; zps = zone périphérique sub-superficielle;
cmc =cellules mères centrales; cm = cordon mérismatique.
Méristèmes radiculaires
Les structures apicales de racines sont aussi variées - sinon plus - que celles des tiges.
A l'extrême pointe de la racine, le pro-méristème est une zone cellulaire de taille réduite où sont situées les initiales méristématiques et quelques unes de leurs dérivées.
Aucune différenciation n'est observable à l'aide des techniques actuelles dans ce groupe de cellules.
Le méristème proprement dit prolonge le pro-méristème vers l'arrière. On choisit de le faire débuter avec le début de la différenciation cellulaire. Il peut se diviser en trois zones concentriques, celles-là même qui se sont organisées dans les tissus de l'embryon y formant trois manchons cylindriques emboîtés l'un dans l'autre :
A la périphérie de l'organe, le protoderme est la première ébauche de ce que sera le tissu de revêtement de la racine, le rhizoderme. C'est également lui qui sera à l'origine de la coiffe.
Au centre, le procambium donnera l'ensemble des tissus du cylindre central, xylème et phloème notamment.
Le méristème fondamental enfin, situé entre protoderme et procambium produira l'écorce de la future racine.
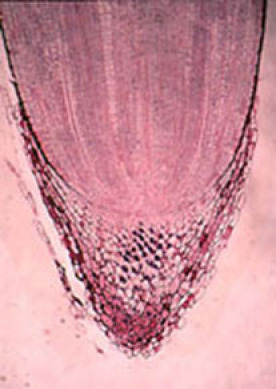
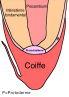
Fig. 04 : Photographie et schématisation d'un méristème racinaire de Zea mais
Au coeur du méristème radiculaire, une zone se distingue par sa faible activité mitotique. Les phases G1 des cycles mitotiques peuvent être très longues (174h).
C'est un centre quiescent de taille réduite (quelques centaines de cellules seulement), qui possède un certain nombre de propriétés remarquables : très grande résistance (à l'irradiation notamment) potentialités méristématiques importantes, etc...