Les angiospermes - Différentiation anatomique
Les cellules d'un végétal vont se différencier, se spécialiser pour former différents tissus. Le regroupement de ces tissus en vue d'assurer les différentes fonctions donneront naissance aux organes : racines, tiges, feuilles et fleurs.
La formation des organes et des tissus résultent de l'activité des méristèmes et a lieu tout au long de la vie de la plante. Ceci est une des caractéristiques des organismes végétaux puisque chez les animaux, la formation des organes et des tissus a surtout lieu durant l'embryogenèse.
Les méristèmes
Ils sont formés de petites cellules indifférenciées isodiamétriques qui se divisent intensément pour assurer le développement de la plante. Les méristèmes primaires apparaissent en premier au cours de l'embryogenèse. Localisés aux extrémités des tiges et des racines, ils assurent la croissance en longueur de la plante et donnent naissance aux tissus primaires et aux méristèmes secondaires. Ces méristèmes secondaires assurent le développement en largeur en augmentant le diamètre des divers axes (tiges, racines). Ils sont à l'origine des tissus dits secondaires. Les méristèmes secondaires existent chez tous les spermaphytes à l'exception des monocotylédones.

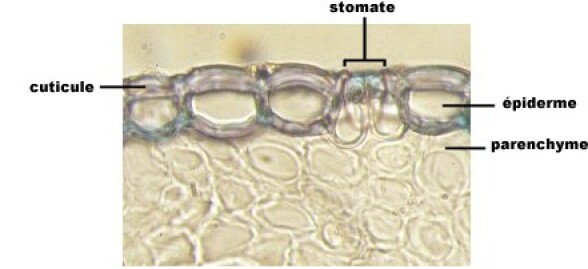
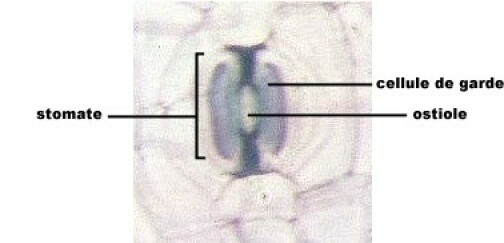
L'épiderme
L'épiderme est le tissu superficiel des feuilles et des tiges jeunes. Il ne comporte, en général, qu'une seule assise cellulaire. La paroi externe des cellules est épaissie d'une couche imperméable, ou cuticule, qui réduit les pertes d'eau par évaporation. L'épiderme est interrompu de place en place par des ouvertures (stomates) bordées par deux cellules stomatiques qui se resserrent ou s'écartent, réglant ainsi les possibilités d'échange gazeux et de vapeur d'eau avec l'extérieur. Dans les plantes à croissance secondaire (gymnospermes et angiospermes dicotylédones), l'épiderme est remplacé par un périderme. Celui-ci provient du fonctionnement d'un méristème secondaire, la zone génératrice subéro-phellodermique, qui produit vers l'extérieur des cellules qui meurent et forment le liège (écorce de tronc d'arbre), vers l'intérieur un tissu vivant, le phelloderme.
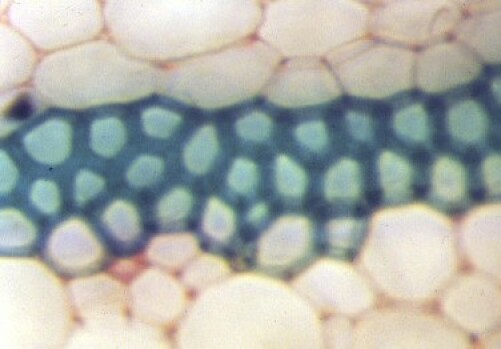
Les tissus vasculaires
Le xylème assure la circulation de la sève brute (sève ascendante), composée d'eau et de sels minéraux puisés par les racines. Il est constitué de cellules mortes dont les parois longitudinales sont épaissies par des dépôts de lignine interrompus de place en place. Chez les fougères, les épaississements sont disposés comme des barreaux d'échelle (trachéides scalariformes) ; chez les gymnospermes, seules de petites formations circulaires, les aréoles, ne sont pas épaissies (trachéides aréolées) ; chez les plantes à fleurs, les épaississements forment des anneaux, une spire continue ou un réseau (trachéides annelées, spiralées, réticulées). Les vaisseaux parfaits, dépourvus de paroi transversale et propres aux plantes à fleurs (angiospermes), sont annelés, spiralés, réticulés, ponctués suivant leur stade de différenciation.


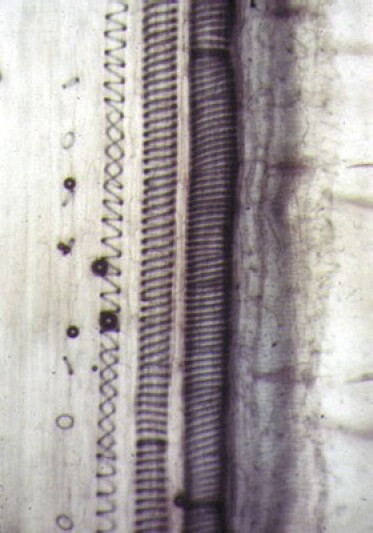
Le phloème assure la circulation de la sève élaborée (sève descendante) qui est enrichie des substances issues de la photosynthèse. Il est composé de tubes criblés constitués de cellules allongées, vivantes mais ayant perdu leur noyau, dont les cloisons transversales sont perforées et au-travers desquelles circule la sève. Ces tubes sont flanqués de cellules compagnes plus petites, vivantes et nucléées, et supposées participer au contrôle des échanges entre tubes criblés et organes végétaux.
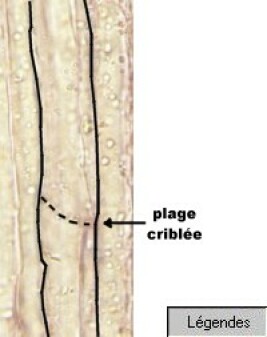
Les éléments de conduction de la sève sont groupés en faisceaux. Dans les racines, faisceaux du xylème et faisceaux du phloème alternent. Dans les tiges, ils sont superposés (ceux du phloème étant les plus externes) en faisceaux indépendants.
Chez les dicotylédones et les gymnospermes, la croissance en diamètre des axes est assurée par un méristème secondaire, le cambium ou zone génératrice libéro-ligneuse, qui se met en place, en une assise continue entre les faisceaux de xylème et de phloème primaires. Ce cambium produit un xylème secondaire, ou bois, vers l'intérieur et un phloème secondaire, ou liber, vers l'extérieur ; l'ensemble de ces productions secondaires formant ainsi un manchon cylindrique. Les plantes à fleurs monocotylédones (graminées, par exemple), dont les axes, le plus souvent, cessent rapidement toute croissance en diamètre, ne possèdent pas de cambium ; de plus, dans une coupe transversale, les faisceaux conducteurs se présentent dispersés dans l'ensemble de la section, alors que chez les dicotylédones ils sont disposés selon un seul cercle.
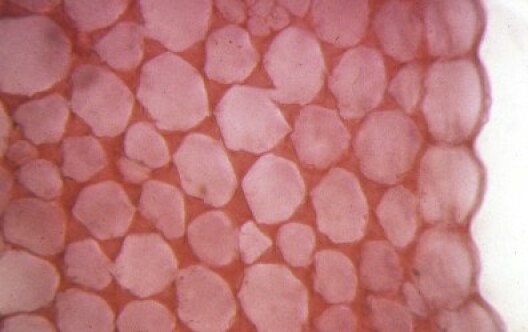
Les tissus assimilateurs et de réserve
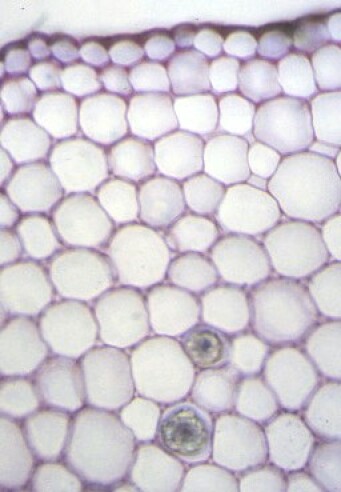
Ce sont les parenchymes nés du fonctionnement des méristèmes. Ils sont formés de cellules vivantes mais qui ne se divisent pas. On distingue les parenchymes assimilateurs, périphériques et chlorophylliens, qui font la photosynthèse, et les parenchymes de réserve, plus internes, qui accumulent des composés organiques (sucres, lipides, protéines). La structure des parenchymes est plus ou moins compacte. Aussi, le parenchyme lacuneux, qui est très poreux, a un rôle dédié aux échanges gazeux avec le milieu.
Les tissus de soutien
A côté des faisceaux de xylème, qui jouent un important rôle de soutien, se trouvent des éléments particuliers formés par des cellules vivantes à parois très épaissies. Ils constituent le collenchyme, aux parois cellulaires cellulosiques, dans les axes jeunes, et le sclérenchyme, aux parois épaissies de lignine et dont les cellules sont mortes, plus abondant dans les végétaux dont la croissance est terminée. Ce type de tissus renforce la résistance mécanique des différents organes de la plante.