Constituants biochimiques
Selon que les cellules sont jeunes ou différenciées, on peut distinguer des parois dites "primaires" et d'autres dites "secondaires".
Ces appellations peuvent être formalisées d'une manière assez simple :
la paroi primaire est la paroi des cellules jeunes en croissance et elle a forcément des propriétés d'extension,
la paroi secondaire est celle des cellules différenciées, que cette différenciation ait affecté chimiquement la paroi (ex. lignification) ou pas.
Les constituants généraux de toutes les parois de spermaphytes sont des polysaccharides, de la cellulose, des pectines et des hémicelluloses ; il s'y ajoute d'autres constituants (des protéines de structure, des protéines enzymatiques, des ions et bien sûr de l'eau).
La cellulose
La cellulose est constituée de polymères de cellobiose (diholoside formé de glucoses liés en \(\beta\)(1-4)). Ces molécules, uniformes sur le plan moléculaire, constituent des fibrilles grâce à des liaisons hydrogènes intra/inter-caténaires. Cette organisation présente une résistance remarquable. La cellulose représente le matériau de base de la paroi cellulaire.
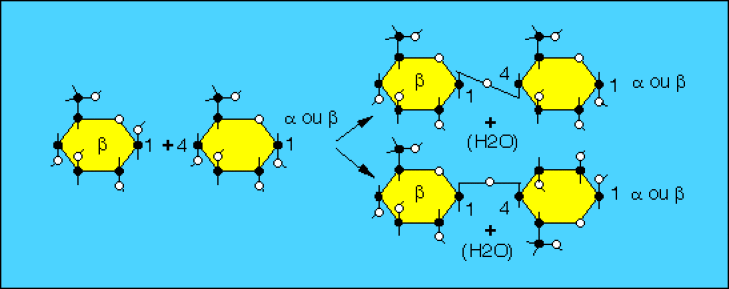
Le motif de base de la cellulose est constitué par le cellobiose. Ce disaccharide (ou diholoside) est formé de deux glucoses liés en \(\beta\)(1-4).
Cette liaison (des glucoses sous forme \(\beta\)) propose deux représentations planes simples. Comme les oxydriles des carbones 1 et 4 sont situés de part et d'autre du plan de la molécule, on peut dessiner la liaison par un trait oblique (en haut) ou inverser la deuxième molécule (en bas). Cette dernière représentation se rapproche le plus de la réalité.
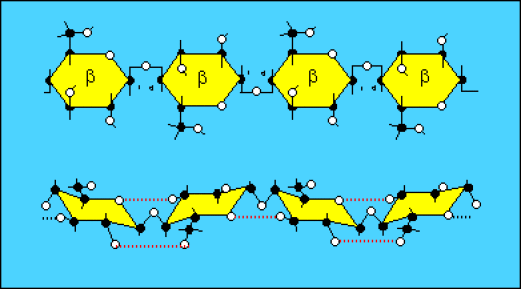
La molécule de cellulose est formée d'un enchaînement de molécules de cellobiose, ce qui revient à un enchainement de molécules de glucose liés en \(\beta\)(1-4). La représentation plane la plus fidèle consiste à retourner un glucose sur deux de 180°. Lorsqu'on représente cette molécule dans une représentation stéréochimique, même simplifiée, on constate :
que cette molécule est monotone (un seul monomère),
que cette molécule est linéaire,
que des liaisons H intra-caténaires peuvent se réaliser de manière répétitive tout au long de la molécule.
Cette molécule linéaire devient alors plus rigide et plus résistante.
Des liaisons H intercaténaires peuvent aussi se réaliser et permettre la formation d'ensembles plurimoléculaires (les microfibrilles) encore plus rigides et résistants.
Pour toutes ces raisons, la cellulose représente la charpente de la paroi cellulaire.
Les pectines
Les pectines représentent une partie importante de la matrice pariétale, c'est à dire de la substance qui englobe les microfibrilles. Ce sont des molécules polysaccharidiques complexes.
En effet, contrairement à la cellulose, elles sont constituées de nombreux monomères différents, associés par des liaisons variées sous forme d'un squelette principalement formé d'acide galacturonique et pourvu de très nombreux branchements. Les propriétés des monomères en font des molécules acides, mais qui peuvent développer localement des propriétés particulières vis à vis des ions (en particulier le calcium).
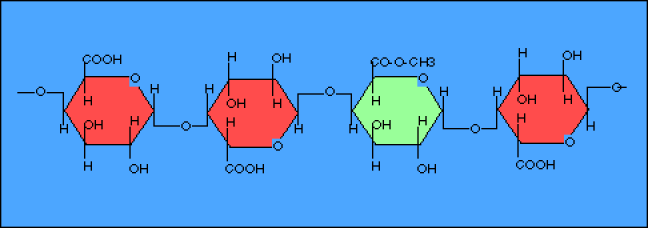
La chaîne principale des pectines est constituée d'un polymère : l'acide polygalacturonique. L'acide galacturonique, qui est le monomère, est un dérivé du galactose (substitution d'une fonction alcool (-OH) par une fonction acide (-COOH) sur le carbone 6. La chaîne est rarement monotone. Certains acides galacturoniques sont méthylés (substitution d'un -H de la fonction acide par une fonction méthyle (-\(CH_{3}\))). Selon le degré de méthylation, l'acide polygalacturonique a des propriétés plus ou moins acides.
La chaîne d'acide polygalacturonique n'est pas monotone ; elle peut être brisée par la présence d'un pentose : le rhamnose.
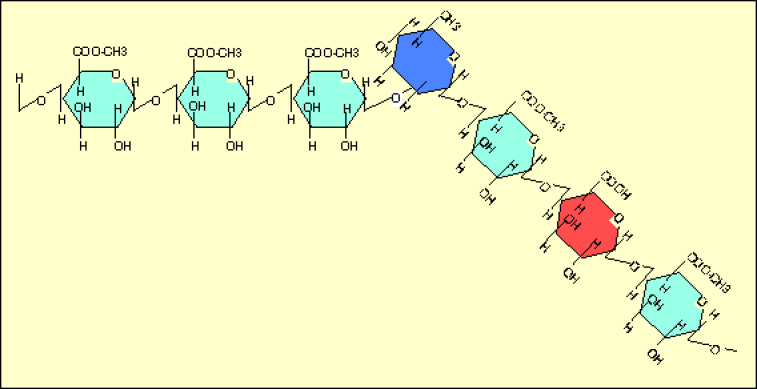
Lorsqu'une région est essentiellement formée d'acide galacturonique non méthylé, des ions calcium divalents peuvent provoquer la jonction de deux chaînes et ainsi amener une sorte de réticulation de la solution de pectines (formation d'un gel).
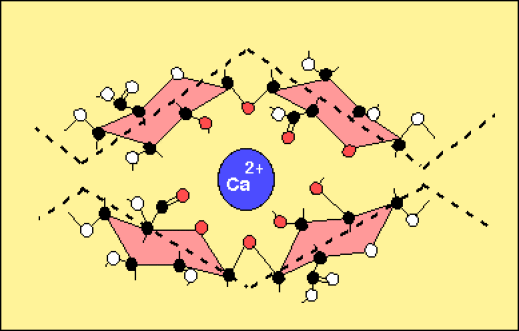
A cet ensemble, il faut ajouter l'addition de nombreuses chaînes latérales constituées par des oses variés (souvent neutres).
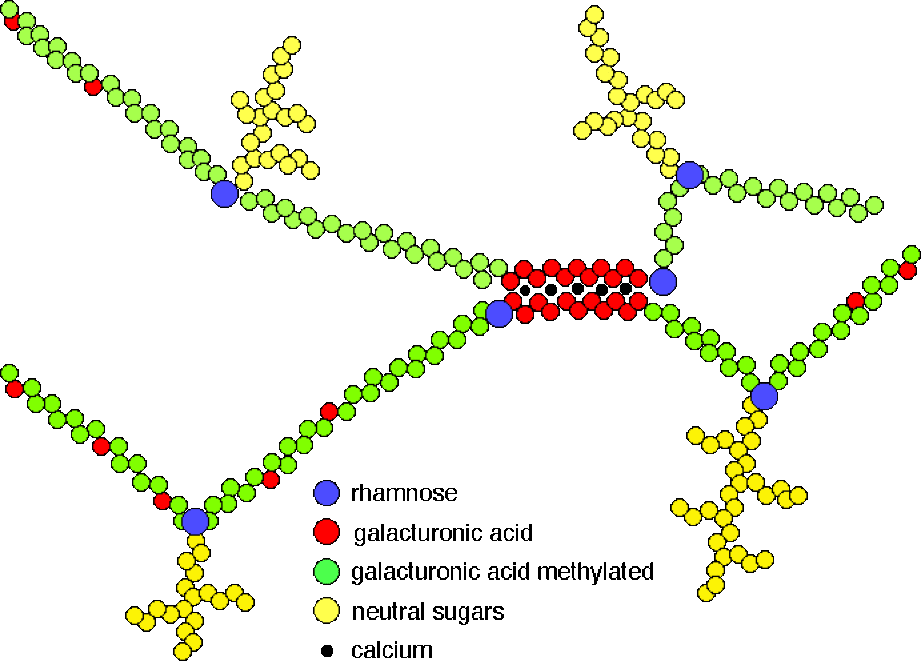
Les pectines représentent des polymères complexes qui varient par la nature de leurs nombreux monomères, leur séquence, la nature et la position des branchements latéraux, etc ...
Les hémicelluloses
Les hémicelluloses représentent une classe de molécules polysaccharidiques très complexe et très variée, car elle comprend de nombreux monomères et chaînes ramifiées.
On commence à connaître certaines séquences, et certaines de ces molécules peuvent avoir une signification informative (en particulier dans la défense vis à vis des pathogènes). Un exemple est étudié, les xyloglucanes, qui peuvent avoir de plus un rôle précis dans la croissance cellulaire.
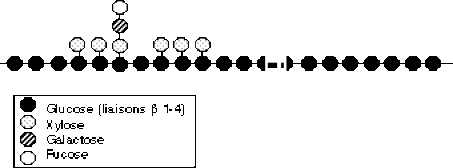
Les xyloglucanes sont formés d'une chaîne de glucose (liés en \(\beta\)(1-4), comme dans la cellulose) sur laquelle se fixent de courtes chaînes latérales de xylose, galactose et fucose.
Remarquons que, par leur structure complexe (séquence des monomères et branchements), les pectines et surtout les hémicelluloses peuvent avoir, de plus, un rôle informatif particulièrement important dans la défense contre les pathogènes.
Les autres constituants et variations
HRGP : Un constituant important sur le plan structural est représenté par certaines protéines particulières.
Il s'agit de protéines riches en un acide aminé particulier : l'hydroxyproline. Dans la cellule différenciée, elles peuvent, par des liaisons covalentes, constituer un réseau avec les autres composants pariétaux et bloquer ainsi les possibilités d'extension de la paroi.
Protéines enzymatiques : Elles sont nombreuses et variées selon les types cellulaires. Les plus communes sont des hydrolases (glucanases), des pectine-methyl-estérases et des peroxydases. Elles doivent jouer des rôles importants dans le comportement biologiques des parois cellulaires.
Des ions : Citons principalement le calcium, qui peut entrer en relation avec les pectines par chélation.