Lois de Mendel (1866)
La génétique classique débute avec les travaux du moine autrichien, Gregor Mendel (1822-1884), qui expérimenta pendant 9 ans sur le Pois (Pisum sativum) pour confirmer ses théories de l'hérédité (démarche utilisée par G. Mendel).
Complément : La démarche expérimentale de G. Mendel.
Gregor Mendel a centré son travail sur un seul organisme (le Pois: Pisum sativum). Il a étudié la transmission de peu de caractères monogéniques mais sur au moins 4 générations.
Le choix du Pois fût très judicieux. En effet :
il existe, pour cette plante, un grand éventail de couples de caractères alternatifs bien définis (couleur et forme des fruits et des graines mûres, couleur des pétales, taille de la plante, etc).
la structure de la fleur permet l'expérimentation :
les étamines et les pistils étant enfermés dans la carène[1] de la fleur, l'autofécondation[2] de la plante est possible.
on peut enlever les étamines avant que le pollen soit mûr et féconder le pistil avec du pollen prélevé sur une autre plante (fécondation croisée).
le temps de génération est court (1 an) et les descendants nombreux.
les hybrides sont aussi fertiles que les parents ce qui permet l'analyse de nombreux descendants sur plusieurs générations.
La démarche expérimentale de G. Mendel peut se décomposer en 5 points :
il a utilisé des lignées pures[3]. Morgan a en effet commencé par multiplier ses plantes par autofécondation pendant 2 ans. Les descendants ainsi obtenus présentaient toujours les mêmes caractères.
le croisement de départ (génération parentale) impliquait des plantes qui ne différaient que par un (monohybridisme[4]) ou deux (dihybridisme) caractères.
il a toujours réalisé des croisements réciproques[5] et des croisements test[6] (test cross[7]).
il a étudié la descendance par autofécondation de chaque hybride sur plusieurs générations.
il a réalisé les mêmes séries d'expériences avec 7 paires de caractères alternatifs.
G. Mendel a publié le résultat de ses études en 1866. À cette époque on ignorait tout de la méiose et des chromosomes mais, en proposant l'existence d'unités héritables (qui seront appelées gènes, en 1906 par le biologiste danois Wilhem Johannsen), Mendel fonda la Génétique. De cette hypothèse Mendel a tiré un certain nombre de principes connus sous le nom de lois de Mendel et applicables à tout eucaryote ayant une méiose normale.
Fondamental : Première loi de Mendel
Les deux allèles d'un gène déterminant un caractère se disjoignent (ségrègent) lors de la formation des gamètes : une moitié des gamètes contient l'un des allèles et l'autre moitié contient l'autre.
Cette loi résulte du fait qu'il y a ségrégation des chromosomes homologues lors de l'anaphase de la première division méiotique (anaphase I).
Impossible d'accéder à la ressource audio ou vidéo à l'adresse :
La ressource n'est plus disponible ou vous n'êtes pas autorisé à y accéder. Veuillez vérifier votre accès puis recharger le média.
Dans des circonstances exceptionnelles où cette loi n'est pas respectée il en résulte habituellement une anomalie dans la descendance.
Fondamental : Seconde loi de Mendel
Si les descendants d'un croisement impliquant deux lignées pures différant par un seul caractère présentent tous le même phénotype, ce phénotype (caractère) est qualifié de dominant (l'autre caractère est dit récessif)
Complément : Limites de la seconde loi de Mendel
1- Les différentes combinaisons alléliques d'un gène rencontrées chez un hétérozygote F1 peuvent donner des phénotypes exactement identiques, différents ou très différents.
Si un hétérozygote A1/A2 présente :
le même phénotype que par exemple l'homozygote A1/A1 : on dit que A1 est dominant et A2 récessif.
Exemple : les caractères du Pois étudiés par G. Mendel.
un phénotype intermédiaire entre ceux des deux homozygotes correspondants : on parle de semidominance, de dominance incomplète, de dominance partielle ou de récessivité incomplète.
Les exemples sont rares chez l'homme mais possibles chez les plantes (ex : la couleur des fleurs).
un phénotype correspondant à la somme des phénotypes des homozygotes correspondants : on parle de codominance.
Exemple : le groupe sanguin MN.
un phénotype plus marqué que celui des homozygotes on parle de superdominance.
Exemple : la vigueur hybride chez les plantes.
2- Les termes utilisés pour décrire les interactions entre allèles conduisant à un phénotype dépendent du mode d'observation de ce phénotype. Exemple : les hémoglobines.
Exemple : Groupe MNSs
En 1927 Karl LANDSTEINER (biologiste d'origine autrichienne alors émigré aux USA) et Philip LEVINE injectent à des lapins des hématies humaines. Les lapins fabriquèrent des anticorps dirigés contre plusieurs antigènes humains voisins. Ils constatèrent que l'un de ces anticorps, appelé anti-M, agglutine les hématies de certaines personnes. Ces hématies sont donc porteuses de l'antigène M. Ils purifièrent également l'anticorps anti-N et découvrirent ainsi le second groupe sanguin appelé groupe MN définit par la présence ou l'absence des antigènes M et/ou N sur les hématies. Contrairement à ce qui se passe pour le groupe ABO, il n'y a normalement jamais les anticorps correspondants dans le sérum. On distingue donc 3 groupes (phénotypes).
Groupe | Antigène présent sur les hématies |
M | antigène M |
N | antigène N |
MN | antigène M et antigène N |

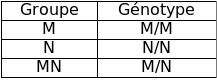
Ce groupe est déterminé par un couple d'allèles, M et N, co-dominants au locus de la glycophorine A sur le chromosome 4 (les 2 glycophorines A codées par ces allèles ne diffèrent que par 2 acides aminés) :
Groupe | Génotype |
M | M/M |
N | N/N |
MN | M/N |
K. LANDSTEINER et Philip LEVINE découvrirent également les sous-groupes S et s. On a montré que ces sous-groupes sont déterminés par 2 allèles codant la glycophorine B (les deux formes de glycophorine B diffèrent par un seul acide aminé). L'allèle S est dominant sur l'allèle s.
On sait maintenant que les deux loci, MN et Ss, sont étroitement liés sur le chromosome n°4 ce qui permet de définir 4 haplotypes possibles : MS, NS, Ms et Ns.
Exemple : Hémoglobines
Chez l'homme on connaît plus de 100 allèles du gène "\(\beta\)" codant la chaîne \(\beta\) de l'hémoglobine. Les plus fréquents sont les allèles \(\beta\)A, \(\beta\)S, \(\beta\)C et \(\beta\)E. L'allèle \(\beta\)A code la chaîne normale, les autres codent des chaînes anormales pouvant conduire à des anémies plus au moins sévères.
Considérons le couple d'allèles \(\beta\)A et \(\beta\)S. Par rapport à l'allèle normal, l'allèle \(\beta\)S présente une mutation ponctuelle dans le codon 146 (changement de A en T) entraînant la formation d'une chaîne \(\beta\)S anormale (changement de l'acide glutamique en valine au niveau de ce codon).
L'hémoglobine contenue dans les hématies est un tétramère constitué de 2 chaînes \(\alpha\) et 2 chaînes \(\beta\) (\(\alpha\)2\(\beta\)2). Le tétramère ne peut se former qu'avec des chaînes \(\beta\) identiques.
Dans les hématies d'un hétérozygote \(\beta\)A/\(\beta\)S, étant donné que les deux allèles s'expriment, on trouve de l'hémoglobine A, HbA (\(\alpha\)2\(\beta\)A2) et de l'hémoglobine S, HbS (\(\alpha\)2\(\beta\)S2) en proportions égales.
Selon le niveau d'analyse du phénotype des hétérozygotes \(\beta\)A/\(\beta\)S par rapport à celui des homozygotes correspondants, on peut tirer les conclusions suivantes :
L'allèle \(\beta\)S est récessif par rapport à \(\beta\)A si on se réfère à l'analyse clinique des sujets.
En effet, seuls les homozygotes \(\beta\)S/\(\beta\)S présentent une anémie falciforme (maladie hémolytique sévère caractérisée par une tendance des hématies à prendre une forme de faucille) nécessitant des hospitalisations notamment à cause d'occlusion aiguë de capillaires, d'infractus de la rate, de déficience splénique etc. Les hétérozygotes sont cliniquement normaux.
L'allèle \(\beta\)S est dominant par rapport à \(\beta\)A si on se réfère au phénotype des hématies observées dans une goutte de sang étalée sur une lame.
Dans ces conditions d'observation, les hématies des hétérozygotes et des homozygotes \(\beta\)S/\(\beta\)S sont en forme de faucille. Cependant dans le sang circulant les hématies des hétérozygotes ont une forme normale.
Les allèles \(\beta\)S et \(\beta\)A sont codominants si s'intéresse à la nature des chaînes \(\beta\) présentes dans les hématies.
Les hématies des hétérozygotes contiennent à la fois de l'HbA et de l'HbS donc des chaînes \(\beta\)A et des chaînes \(\beta\)S car les deux allèles s'expriment également.
Les allèles \(\beta\)S et \(\beta\)A sont semidominants si on analyse les individus en atmosphères peu oxygénées (dans un avion non pressurisé en haute altitude par exemple).
En effet, le phénotype des \(\beta\)A/\(\beta\)S alors intermédiaire entre ceux des homozygotes. Dans ces conditions les homozygotes \(\beta\)A/\(\beta\)A n'ont pas de problème (leurs hématies sont de forme normale) mais toutes les hématies des homozygotes \(\beta\)S/\(\beta\)S prennent la forme de faucille qui les rend moins flexibles : elles ne peuvent plus passer dans les petites capillaires ce qui entraîne des arrêts du flux sanguin causant des hypoxies locales. Les hétérozygotes auront des anomalies de circulation sanguine moins graves car seuls quelques pourcents de leurs hématies prennent la forme de faucille.
Le phénotype des \(\beta\)A/\(\beta\)S est superdominant si on considère la réponse à la malaria.
En effet les hétérozygotes sont résistants à cette maladie alors que les deux types d'homozygotes y sont sensibles.
Fondamental : Troisième loi de Mendel
La ségrégation d'un couple d'allèles est indépendante de celle d'un autre couple d'allèles.
Complément : Limites de la troisième loi de Mendel
Nous savons maintenant que cette loi n'est vraie que si les deux couples d'allèles sont sur des chromosomes non homologues ou très loin l'un de l'autre sur le même chromosome.