Ségrégation d'un couple d'allèles
Explication :
Prenons un exemple et observons les asques formés à la suite d'un croisement entre deux souches de Neurospora de signe conjuguant compatible.
Une des souches parentales est une souche sauvage de référence, dont les spores sont pigmentées en brun foncé (disons noir, pour simplifier). L'autre parent est une souche mutante dont les spores, dépourvues de pigment, sont blanches.

Tout d'abord on observe, qu'au-delà des différences (voir ci-dessous), les asques formés présentent tous les points communs suivants :
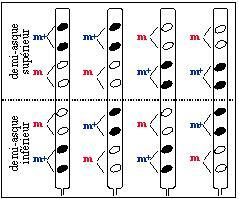
quel que soit leur ordonnancement, les asques contiennent tous quatre spores noires et quatre spores blanches, il s'agit de la ségrégation de deux allèles d'un même gène, nous allons l'illustrer tout de suite ;
si on part d'une extrémité de l'asque, les spores prises deux à deux sont toujours identiques, cela n'est pas étonnant puisque chaque paire provient par mitose d'une cellule.
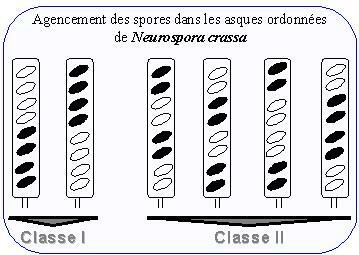
La seconde observation concerne l'ordre des spores dans les asques : sur ce critère il y a six catégories d'asques que l'on peut regrouper en deux classes. Dans la classe I, les demi-asques sont homogènes quant au phénotype des spores qu'ils contiennent. Dans la classe II, les demi-asques sont hétérogènes.
Comment expliquer cette observation ?
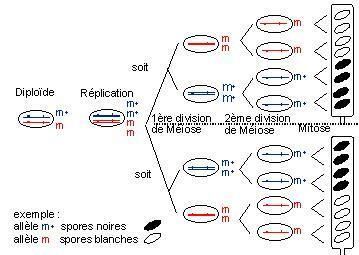
Voyons ce qui se passe lors de la méiose si l'on considère un gène présent sous deux formes alléliques différentes chez les souches parentales. Appelons m l'allèle mutant et m+ l'allèle sauvage. Au moment de la prophase de la première division de la méiose, les chromosomes homologues qui portent chacun de ces allèles s'apparient. Ils sont dupliqués sur toute leur longueur, sauf au niveau du centromère. Des crossing-over, des échanges de chromatides s'effectuent à divers endroits au cours de cet appariement.
Par rapport au locus qui nous intéresse on distingue a priori deux cas alternatifs :
soit il n'y a pas de crossing-over entre le locus du gène et le centromère du chromosome qui le porte,
soit il y a un ou plusieurs crossing-over entre le locus du gène et le centromère du chromosome qui le porte
En conclusion :
La ségrégation d'un couple d'allèles à chaque méiose se fait dans les proportions 1/2-1/2.
Les premières observations de ce type ont apporté la confirmation expérimentale de ce que l'on appelait la théorie chromosomique de l'hérédité, à savoir que le comportement des gènes se calquait sur celui des chromosomes au moment des divisions cellulaires.
Pour tout croisement impliquant un couple d'allèles on doit trouver des quantités statistiquement égales des deux catégories d'asques qui correspondent à la ségrégation des allèles à la méiose I. Les quatre autres types d'asques, ceux qui correspondent à la ségrégation des allèles à la méiose II, doivent aussi se trouver en proportions égales.
Pour aller plus loin, voir la partie "Apports de l'analyse en tétrades" du chapitre "Ségrégation de deux couples d'allèles". Il y est traité de la localisation du centromère d'un chromosome sur la base du pourcentage des asques de la classe II. Ce pourcentage est, en effet, corrélé à la probabilité qu'il se produise un ou plusieurs crossing-over entre le locus considéré et le centromère du chromosome, autrement dit à la probabilité de recombinaison entre ces deux marqueurs et donc finalement à la distance qui les sépare.
Exemple : aucun crossing-over entre le locus du gène et le centromère.
Dès la première division de la méiose, il y a séparation, ou ségrégation, dans deux noyaux différents, des deux allèles du gène considéré. On dit qu'il y a eu ségrégation à la première division de la méiose (à la méiose I).
Par ailleurs, l'un ou l'autre des centromères migre avec une probabilité égale vers l'une ou l'autre des extrémités du futur asque. Ce type d'événement est à l'origine des asques de la classe I dont les demi-asques sont homogènes. Les deux types d'asques de la classe I doivent apparaître en nombre statistiquement égal.
Exemple :
un crossing-over entre le locus du gène et le centromère, l'échange de chromatides au cours de la première division méiotique conduit à deux noyaux qui, cette fois-ci, contiennent encore les deux allèles m et m+ du gène. De plus, la migration des centromères vers l'un ou l'autre pôle de la cellule se fait toujours au hasard.
Ce n'est qu'au cours de la deuxième division méiotique que les deux allèles se sépareront, avec encore une fois une probabilité égale pour chacun de migrer vers le haut ou vers le bas du futur asque ordonné.
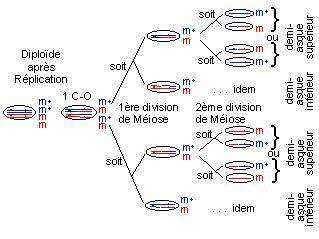
On parle de ségrégation à la deuxième division de la méiose (à la méiose II).
C'est ainsi que l'on peut expliquer la formation des quatre catégories d'asques de la classe II, à demi-asques hétérogènes, qui doivent être équiprobables.